The Blood Test That Should Have Shown NAD+ Declining With Age. It Did Not.
Prefer to listen? Hit play for a conversational, audio‑style summary of this article’s key points.
NAD+ is a coenzyme found in every cell in the body and plays a central role in energy production, DNA repair, sirtuin activation, and cellular stress response. Its decline in animal tissues with age generated substantial excitement about the possibility that restoring NAD+ through supplementation with precursors like NR and NMN could slow aging. That animal data remains real and the biological mechanisms are well characterized.
A new study published in Nature Metabolism subjected the foundational assumption of age-related blood NAD+ decline to the most rigorous human testing yet conducted. Using a validated measurement method applied across seven independent human cohorts totalling more than 300 individuals, the researchers found that whole-blood NAD+ levels were remarkably stable across age and lifestyle interventions.
Blood NAD+ did not decline with age in any cohort examined. Younger adults and older adults showed no significant difference. NAD+ plotted continuously against age across cohorts spanning the twenties through the late eighties showed essentially flat regression lines, with age accounting for approximately one percent of the variation in NAD+ levels.
Elite athletes showed the same blood NAD+ levels as sedentary controls. If sustained extreme physical training was going to reveal differences in NAD+ metabolism, this was the population in which to look. No difference was detected, although some NAD+-related metabolites showed modest differences in athletes.
Exercise and nutritional supplementation did not move blood NAD+ in frail older adults. A three month resistance exercise intervention, a protein-rich dietary intervention, a six month multimodal exercise and nutrition intervention: none of these produced measurable changes in whole-blood NAD+ concentration.
NR supplementation did raise blood NAD+ as expected, confirming the measurement method was sensitive enough to detect genuine changes when they occurred. This is a critical interpretive point. The absence of change across the other six cohorts was not a consequence of an insensitive assay. The method could see the signal when the signal was there.
The earlier studies reporting age-related NAD+ decline in human blood may have been detecting a measurement artifact rather than a biological reality. Freezing blood samples before measurement causes NAD+ to degrade by 21 to 27 percent through a specific mechanism involving red blood cell membrane rupture and exposure to NAD+-consuming enzymes including CD38. This degradation is variable across samples, meaning it can create apparent biological differences where none exist.
Many prior studies were likely underpowered to detect genuine age-related NAD+ differences even if those differences were real. Under the variance structure typical of studies that include a single freeze-thaw cycle, cohorts of 20 individuals per group can only reliably detect differences larger than 7 nanomoles per milliliter. Several studies reporting age-related declines used smaller cohorts and reported effect sizes below this threshold.
This study does not say NAD+ is unimportant for aging biology or that NR and NMN supplementation produces no biological effects. It says that whole-blood NAD+ is not a reliable biomarker of biological aging in humans, and that using blood NAD+ as a readout of whether supplementation is engaging aging biology may be measuring the wrong thing. What happens in blood may not reflect what is happening in muscle, liver, brain, or other tissues where NAD+ metabolism matters most.
Introduction: Why NAD+ Became One of Longevity Science's Most Discussed Molecules
Few molecules have generated as much excitement in longevity research over the past decade as nicotinamide adenine dinucleotide, better known as NAD+. It has become the biological rationale behind a multibillion dollar supplement industry, the subject of hundreds of scientific papers, and the centerpiece of a compelling narrative about why we age and what might be done about it. Understanding why that narrative took hold requires understanding what NAD+ actually does, and why its apparent decline with age seemed so significant.
NAD+ is not exotic. It is a coenzyme found in every cell in the body, as fundamental to cellular biology as any molecule in the human metabolome. Its primary role is to shuttle electrons during the chemical reactions that convert food into usable energy, a process that occurs continuously in the mitochondria of every metabolically active cell. Without NAD+, this energy conversion process cannot function. Without adequate energy conversion, cells cannot do their jobs. And when cells cannot do their jobs across enough tissues and organs, the consequences look a great deal like aging.
But NAD+'s significance extends beyond energy metabolism. It is also a required substrate for a class of proteins called sirtuins, which regulate gene expression, coordinate cellular stress responses, and have been repeatedly associated with longevity in model organisms. It fuels the PARP enzymes responsible for detecting and repairing DNA damage. It supports a network of cellular maintenance processes that, when functioning well, keep tissues healthy and resilient, and when compromised, allow the accumulated damage that drives aging to proceed unchecked.
The excitement began in earnest when studies in animals, primarily mice, showed that NAD+ levels in tissues declined substantially with age, and that restoring those levels through supplementation with NAD+ precursors produced striking effects: improved mitochondrial function, enhanced physical performance, and in some studies, extended lifespan. The biological story was coherent, the animal data was compelling, and the commercial opportunity was obvious. Within a few years, supplements containing nicotinamide riboside and nicotinamide mononucleotide, two precursor molecules the body can convert into NAD+, had become among the most widely purchased longevity products on the market.
Underlying all of this was a foundational assumption: that NAD+ levels in human blood decline with age, just as they appeared to do in animal tissues, and that this decline could be detected through blood testing and reversed through supplementation. It was an assumption that seemed well supported, at least initially. Several human studies reported age-related declines in blood NAD+ levels. The narrative cohered. The supplements sold.
A new study published in Nature Metabolism has now subjected that assumption to the most rigorous human testing it has yet received. The results were not what the field expected.
What NAD+ Is and Why It Matters
To understand why the new findings are significant, it helps to start with what NAD+ actually does inside a cell, and why the possibility of its decline with age captured the attention of researchers and the supplement industry alike.
Think of NAD+ as a molecular shuttle bus. Its job is to pick up electrons generated during the breakdown of food and carry them to the mitochondria, where those electrons are used to power the production of ATP, the energy currency that cells run on. This process, called cellular respiration, is happening continuously in every metabolically active tissue in the body. The brain, the heart, the muscles, the liver: all of them depend on a reliable supply of NAD+ to keep energy production running efficiently. When NAD+ is abundant, this system operates smoothly. When it is depleted, the entire downstream machinery of energy production begins to falter.
But NAD+ does considerably more than shuttle electrons. It is also the required fuel for a family of proteins called sirtuins, which function as master regulators of cellular health. Sirtuins control how genes are expressed in response to stress, coordinate the repair of damaged DNA, regulate the inflammatory signals that accumulate with age, and help determine whether a cell responds to metabolic stress by adapting and surviving or deteriorating and dying. In model organisms from yeast to mice, activating sirtuins has consistently been associated with extended lifespan and improved metabolic resilience. Because sirtuins consume NAD+ to do their work, anything that reduces NAD+ availability effectively throttles their activity.
NAD+ is also the substrate required by PARP enzymes, which patrol the genome for DNA damage and orchestrate its repair. DNA damage accumulates continuously as a byproduct of normal cellular metabolism, and the efficiency with which it is repaired is one of the primary determinants of how quickly cells age and malfunction. PARP enzymes are voracious consumers of NAD+, particularly under conditions of high oxidative stress, which means that when NAD+ is scarce and DNA damage is frequent, the system faces a compounding problem: the very conditions that increase demand for NAD+-dependent repair are the conditions most likely to deplete the NAD+ supply.
Taken together, these roles position NAD+ at the intersection of several of the most important biological processes implicated in aging: energy metabolism, gene regulation, DNA repair, and cellular stress response. It is not an exaggeration to describe it as a central node in the cellular maintenance machinery that healthy aging depends on.
The excitement generated by animal studies showing NAD+ decline with age was therefore biologically coherent. If tissue NAD+ falls as animals age, and if that fall compromises the sirtuins, the PARP enzymes, and the mitochondrial energy production that cells depend on, then restoring NAD+ through supplementation might meaningfully slow the biological processes driving aging. The precursor molecules nicotinamide riboside and nicotinamide mononucleotide offered a practical route to doing this: both can be taken orally, both are converted by the body into NAD+, and both were shown in animal studies to raise tissue NAD+ levels and produce the kinds of functional improvements the hypothesis predicted.
What remained less certain, even as the supplements proliferated and the market expanded, was whether the same decline was occurring in humans, whether blood NAD+ was tracking it reliably, and whether supplementation was doing in human tissues what it appeared to do in mice. These were not small uncertainties. They were the questions on which the entire clinical and commercial rationale for NAD+ supplementation ultimately rested. And they were the questions the new study set out to answer.
NAD+ is at the intersection of several of the most important biological processes implicated in aging: energy metabolism, gene regulation, DNA repair, and cellular stress response. It is not an exaggeration to describe it as a central node in the cellular maintenance machinery that healthy aging depends on.
The Problem With the Evidence So Far
Before the new study was conducted, the human evidence for age-related NAD+ decline in blood was in an uncomfortable state: widely cited, commercially influential, and scientifically inconsistent.
Some studies had reported meaningful associations between age and blood NAD+ levels, finding lower concentrations in older adults and interpreting this as evidence that the age-related decline observed in animal tissues was also occurring in humans. These findings were incorporated into the rationale for NAD+ supplementation and cited repeatedly in the marketing materials of supplement companies. The narrative they supported was intuitive: NAD+ falls with age in both animals and humans, supplementation can restore it, and restoring it should produce the benefits observed in animal studies.
Other studies told a different story. Several independent research groups measuring NAD+ in human blood found no clear association with age in unadjusted analyses. The findings did not align, and the disagreement was not trivial. When independent laboratories measuring the same molecule in similar populations arrive at contradictory conclusions, the explanation is usually one of three things: genuine biological variability between study populations, different analytical methods producing different results, or pre-analytical handling differences corrupting the samples before measurement even begins.
The third possibility turned out to be the most important one, and it points to a problem that is easy to overlook but consequential in its implications.
NAD+ is not a stable molecule once it leaves the body. It is predominantly located inside cells, particularly red blood cells, and it is surrounded in the bloodstream by enzymes whose job is to consume it. Two of these enzymes, CD38 and CD157, are particularly relevant. They are present on the surface of red blood cells and other immune cells, and they are efficient NAD+-degrading machines. Inside intact, living cells, NAD+ is protected from these enzymes by the cell membrane. But when blood is frozen and the cell membranes begin to lyse, that protection disappears. The NAD+ that had been safely sequestered inside cells is suddenly exposed to the very enzymes designed to break it down.
The practical consequence of this biology is that freezing a blood sample and thawing it before measuring NAD+ can reduce the apparent NAD+ concentration by 21 to 42 percent, depending on how many freeze-thaw cycles the sample has undergone. This is not a small analytical error. It is a degradation large enough to look like a biologically meaningful decline, and it is highly variable across samples, meaning it introduces noise into measurements that can masquerade as a real signal.
The critical implication is this: studies that collected blood samples, froze them for storage, and later thawed them for measurement may have been detecting the consequences of sample handling rather than the biological reality of what was in the blood when it was drawn. If older samples happened to be stored differently, handled more frequently, or subjected to more freeze-thaw cycles than younger samples, the resulting apparent age-related difference in NAD+ could be entirely artifactual. The biology would look real. The statistics might even be significant. But the signal would be a product of the laboratory rather than the body.
This was the problem the Amsterdam UMC team set out to solve. Rather than simply measuring NAD+ in more people, they first built and validated a measurement system rigorous enough to distinguish genuine biological signal from the handling-induced noise that had been contaminating the field's conclusions. The approach they took, and what it revealed when applied across seven independent human cohorts, is what makes this study different from everything that preceded it.
The Study: Seven Cohorts, One Rigorous Method
The first thing the Amsterdam UMC team did was not measure NAD+ in people. It was figure out how to measure NAD+ in people correctly.
This sequencing matters. Before any biological conclusions could be drawn from blood NAD+ measurements, the team needed to understand precisely how much their measurement system was contributing to the numbers it produced, and how much of that contribution could be mistaken for biology. This required building and validating a measurement method from the ground up, characterizing every major source of variability that could corrupt a NAD+ measurement before the data from human cohorts was interpreted.
The method they developed uses ultra-high-performance liquid chromatography coupled with high-resolution mass spectrometry, a highly sensitive analytical platform capable of detecting and quantifying NAD+ at the concentrations present in just ten microliters of whole blood, roughly a fifth of a standard droplet.
To account for the variability introduced during sample processing, they incorporated a stable isotope-labeled internal standard, a chemically identical but isotopically distinct version of NAD+ that behaves identically to the molecule being measured throughout the extraction and analysis process.
By tracking how the internal standard moves through the system, the team could correct for procedural losses and ensure that what they were measuring reflected what was actually in the sample rather than what survived the analytical process.
The validation work that followed was methodical and revealing. The team measured the method's sensitivity, establishing the lowest concentration of NAD+ it could reliably detect and quantify. They tested its reproducibility by running the same sample ten times within a single analytical session and across ten separate sessions on different days, establishing how much variation the method itself introduced. Both figures were within acceptable ranges for a rigorous quantitative assay. They confirmed that the measurement scaled linearly with increasing blood volume, meaning the numbers it produced were not distorted by the amount of sample used. And they verified accuracy through spike-recovery experiments, spiking samples with known amounts of NAD+ and confirming that the method recovered them at expected concentrations.
Then came the handling experiments, which turned out to be as important as anything else in the paper. The team systematically tested what happened to NAD+ under the storage and processing conditions commonly used in clinical and research settings. The findings were striking. Freezing whole blood at minus 20 degrees Celsius caused a 27 percent reduction in measured NAD+. Freezing at minus 80 degrees caused a 21 percent reduction. Repeated freeze-thaw cycles compounded the loss progressively, with each cycle adding further degradation. Dried blood spots, another common collection method, showed degradation comparable to frozen small-volume samples. The only conditions that reliably preserved NAD+ were methanol-based stabilization methods applied either before freezing or, in a practical compromise, added to the frozen sample before thawing.
Having characterized the handling problem precisely, the team then performed the power calculations necessary to understand what cohort sizes would be needed to detect genuine biological differences in NAD+ under real-world conditions. The answer was sobering. Under the variance structure typical of studies that include a single freeze-thaw cycle, a cohort of 20 individuals per group, which matched their cross-sectional aging cohort, could reliably detect differences only of approximately 7 nanomoles per milliliter or larger. Many of the age-related differences reported in prior human studies fell below this threshold, meaning those studies were almost certainly underpowered to detect the effects they claimed to have found even if those effects were real.
With the method validated, the handling effects quantified, and the statistical power requirements understood, the team applied their approach across seven independent human cohorts with distinct designs.
- The cross-sectional aging cohort compared younger adults under 30 with older adults over 60. The CardioHT cohort examined NAD+ across a continuous age range in adults with elevated cardiovascular risk.
- The ELITE cohort included elite athletes, younger non-athlete controls, and older non-athlete controls, providing a test of whether extreme physical activity levels revealed differences in NAD+ metabolism.
- The Leiden Longevity Study cohort examined 70 older adults between the ages of 63 and 87, providing a window into NAD+ stability across later decades of life.
- The TEAMS cohort tested whether a three month exercise intervention, with or without a protein-rich diet, changed whole-blood NAD+ in frail older adults.
- The MEJNES2019 cohort tested whether a six month multimodal intervention combining nutritional supplementation and exercise produced changes in frail older adults.
- And the twin-pair NR supplementation cohort, in which participants took escalating doses of nicotinamide riboside over five months, served as the positive control: if the method was sensitive enough to detect genuine changes in NAD+ when a direct precursor was administered, it should show an increase here.
Together these cohorts covered a wide range of ages, fitness levels, health statuses, and intervention types. If whole-blood NAD+ was going to reveal age-related decline or respond to lifestyle interventions anywhere, it had every opportunity to do so across this range of human contexts. What the data showed across all of them is the finding that gives this paper its significance.
What the Data Showed
The results across six of the seven cohorts were consistent in a way that is unusual in human biology research, where noise, variability, and contradictory findings are the norm. Across every population examined, using a measurement method specifically designed to distinguish genuine biological signal from handling artifacts, whole-blood NAD+ levels were remarkably stable.
In the cross-sectional aging cohort, younger adults under 30 and older adults over 60 showed no significant difference in whole-blood NAD+ concentrations. This was the most direct test of the age-decline hypothesis the study offered, matching two groups specifically chosen to maximize the likelihood of detecting an age-related difference if one existed. It did not exist, at least not at a magnitude detectable in a well-powered cohort using a rigorously validated method.
The CardioHT cohort extended this finding across a continuous age range. When NAD+ levels were plotted against age for 26 adults spanning from their late twenties to their early seventies, the regression line was essentially flat. The R-squared value, a statistical measure of how much of the variation in NAD+ could be explained by age, was 0.012. In practical terms, this means age accounted for approximately one percent of the variation in NAD+ levels across this cohort. The remaining 99 percent was attributable to other sources of variability, none of which appeared to be systematically related to how old the participants were.
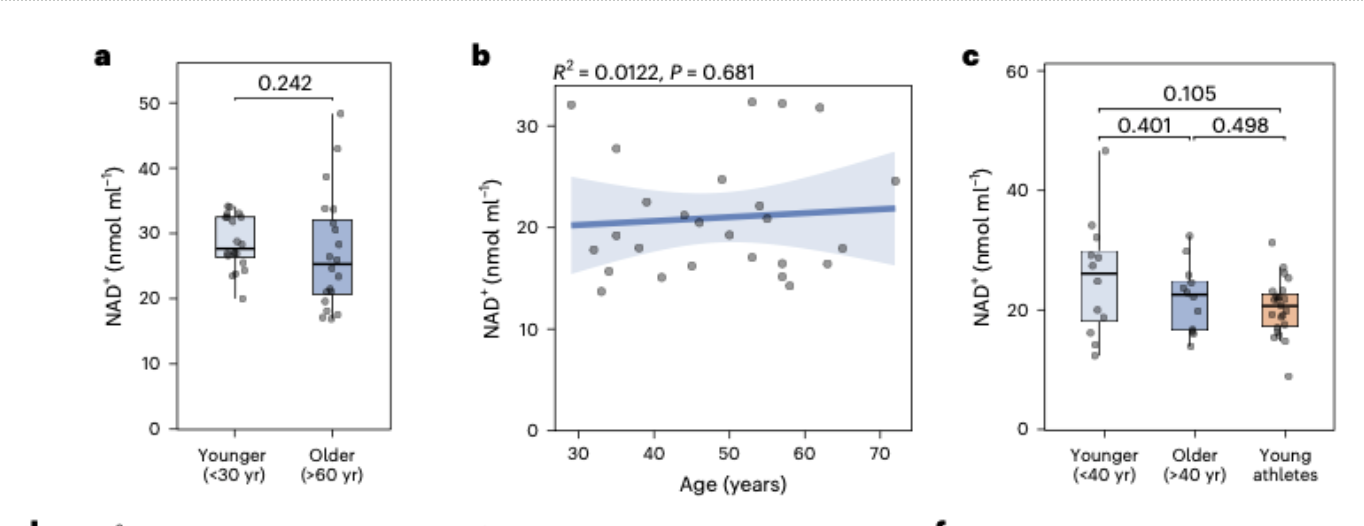
The ELITE cohort produced a similarly unremarkable finding from what might have been expected to be a revealing comparison. Elite athletes competing at national and international levels, whose bodies represent a physiological extreme in terms of metabolic demand and cardiovascular adaptation, showed NAD+ levels comparable to younger non-athlete controls. Older non-athlete controls were similarly indistinguishable from their younger counterparts. If sustained intense physical training was going to reveal differences in NAD+ metabolism, this was the population in which to look. The differences were not there. Some NAD+-related metabolites showed modest differences in athletes, suggesting that the broader NAD+ metabolome is not entirely unresponsive to extreme exercise, but the NAD+ concentration itself remained stable.
The Leiden Longevity Study cohort narrowed the lens to later life specifically, examining 70 individuals between the ages of 63 and 87. This age range is where the consequences of biological aging are most apparent and where one might most expect to see the metabolic deterioration that a declining NAD+ supply would theoretically drive. The regression of NAD+ against age across this 24-year span produced an R-squared of 0.051 and a p-value of 0.991. NAD+ levels were stable across the full age range examined, with no detectable trend in either direction.
The intervention cohorts added a further dimension to the picture. In the TEAMS cohort, frail older adults who completed a three-month supervised resistance exercise program showed no change in whole-blood NAD+ relative to baseline. The same was true for the group that combined exercise with a protein-rich diet targeting 1.2 to 1.5 grams of protein per kilogram of body weight per day. Exercise, even structured resistance training in a population that might be expected to show metabolic responsiveness, did not move blood NAD+.
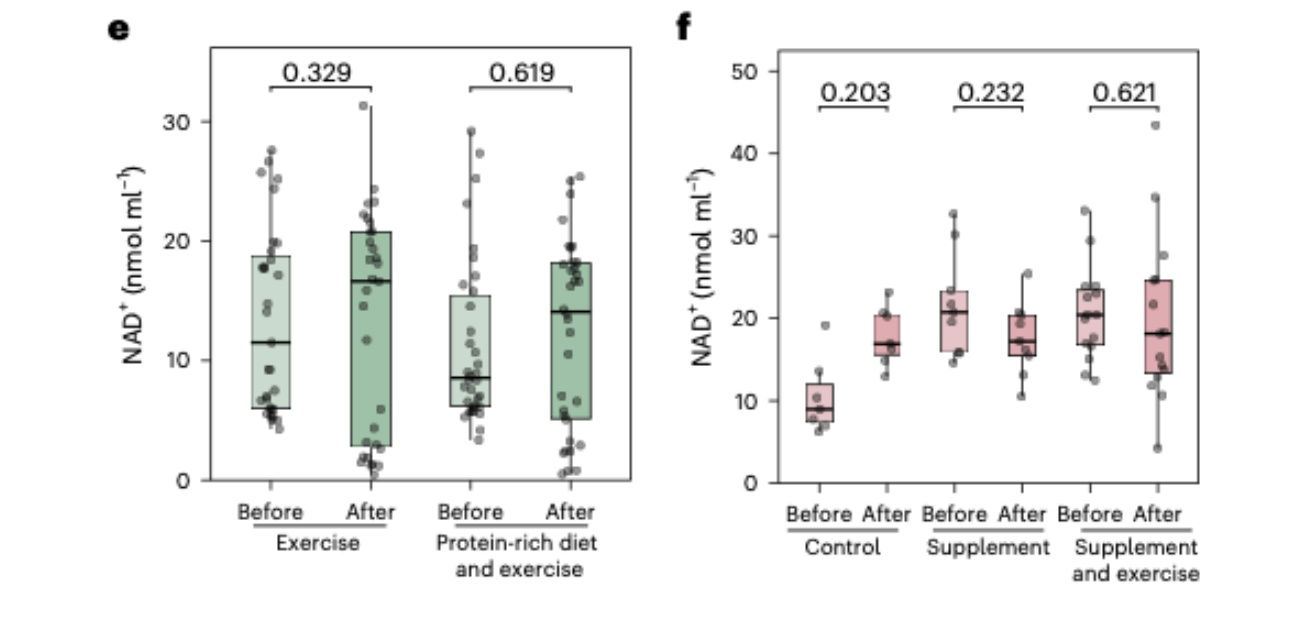
The MEJNES2019 cohort, which tested a six-month multimodal intervention in frail older adults combining nutritional supplementation and exercise, produced the same result. Control, supplementation alone, and supplementation combined with exercise: NAD+ remained unchanged across all three groups over the six-month study period. Some NAD+-related metabolites showed modest shifts in certain intervention groups, suggesting that the broader biochemical landscape of NAD+ metabolism is not entirely static, but the NAD+ concentration itself did not move.
Then came the result that confirmed the method was working as intended. In the twin-pair NR supplementation cohort, participants who took escalating doses of nicotinamide riboside over five months, reaching one gram per day, showed a clear and statistically significant increase in whole-blood NAD+. This was the expected result, consistent with multiple prior studies showing that NR supplementation raises blood NAD+ levels. Its presence in the dataset serves a critical interpretive function. It establishes that the measurement system was sensitive enough to detect genuine changes in NAD+ when they occurred, and that the absence of changes across the other six cohorts was not a consequence of an insensitive assay failing to see real effects. The method could see the signal when the signal was there. In six of the seven cohorts, the signal was not there.
The team also assessed two composite measures of NAD+ metabolism, the niacin index and the niacin number, which reflect the ratio of NAD+ to related metabolites and are used as indicators of overall niacin nutritional status. Neither measure showed substantial changes across any of the cohorts, adding further weight to the conclusion that the stability observed in NAD+ itself was not masking compensatory shifts in the broader metabolic network.
Taken together, the data from these seven cohorts, spanning young and old adults, elite athletes and frail older adults, sedentary individuals and those undergoing structured exercise and nutritional interventions, told a coherent and unexpected story. Whole-blood NAD+ in humans is not a sensitive readout of aging. It does not decline with age in a pattern that routine blood testing can detect. It does not respond to the lifestyle interventions most commonly recommended alongside NAD+ supplementation. And it does respond, clearly and predictably, to direct precursor supplementation, confirming that the molecule is not biologically inert but that blood levels simply do not reflect the age-related dynamics the supplement narrative assumed they would.
Why the Earlier Studies May Have Been Wrong
The consistency of the new findings across seven independent cohorts raises an uncomfortable question. If whole-blood NAD+ does not decline with age under rigorous measurement conditions, why did earlier studies report that it did?
The answer, as the Amsterdam UMC team's handling experiments reveal, is likely methodological rather than biological. And understanding precisely how sample handling corrupts NAD+ measurements is essential for evaluating not just the prior literature on age-related NAD+ decline but the broader field of NAD+ biomarker research.
The core problem is the freeze-thaw cycle. In standard research and clinical practice, blood samples are collected, placed in tubes, and frozen for storage, often at minus 80 degrees Celsius, until the laboratory is ready to process them. This is a routine and generally sensible approach to sample preservation. For most biomarkers, freezing introduces manageable and relatively uniform degradation that can be accounted for in the analysis. For NAD+, the situation is considerably more complicated.
NAD+ is predominantly located inside red blood cells, where it is protected from the extracellular environment by the intact cell membrane. Circulating in the plasma outside those cells are enzymes, particularly CD38 and CD157, whose biological function includes breaking down NAD+ and related molecules. Under normal physiological conditions, these enzymes have limited access to the intracellular NAD+ pool because the cell membrane keeps them separated. When blood is frozen, however, red blood cell membranes begin to rupture through a process called hemolysis, releasing their intracellular contents into the surrounding plasma. The NAD+ that had been safely contained inside the cells is suddenly exposed to the very enzymes designed to consume it.
The result, as the Amsterdam UMC team measured directly, is substantial and variable NAD+ loss. A single freeze-thaw cycle at minus 20 degrees caused a 27 percent reduction in measured NAD+ concentration. At minus 80 degrees, the reduction was 21 percent. Each additional freeze-thaw cycle compounded the loss further. And critically, this degradation was not uniform across samples. The standard deviation of the difference between fresh and frozen samples was 3.5 nanomoles per milliliter, meaning that two samples with identical true NAD+ concentrations could show apparently different concentrations after freezing simply because the extent of membrane rupture and enzymatic degradation varied between them.
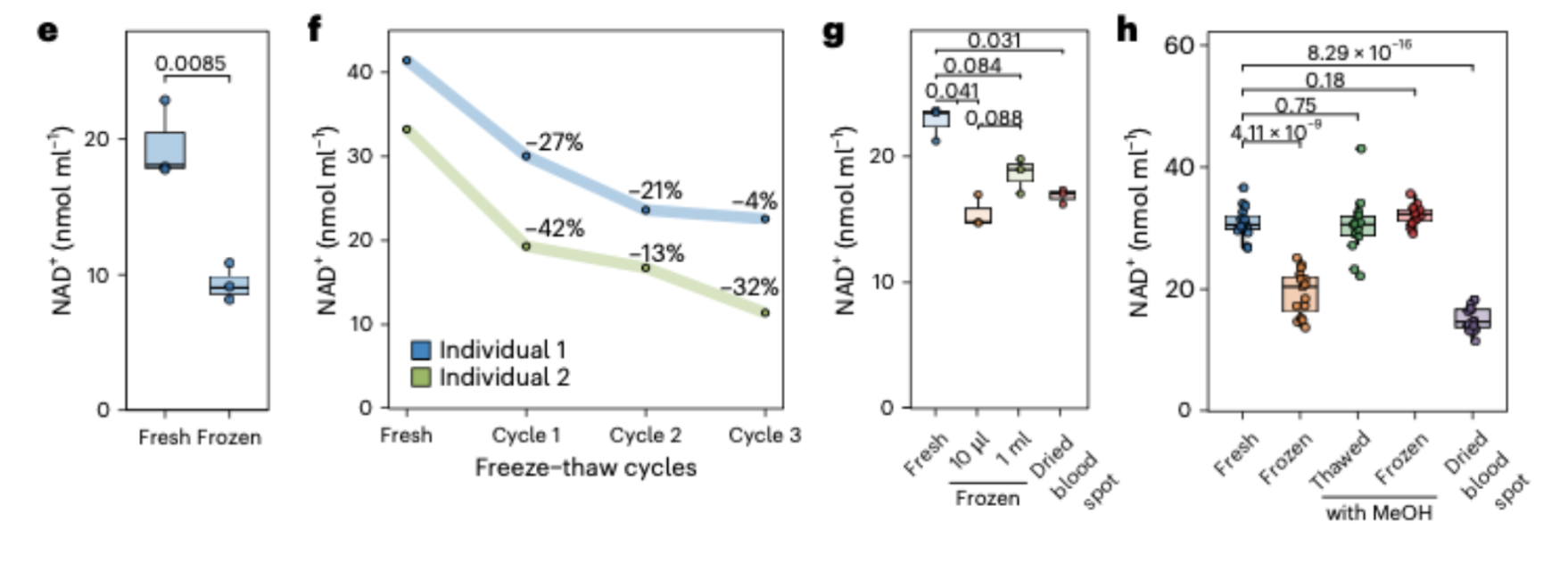
This variability is what transforms a handling artifact into something that can look like a biological signal. If an older study collected blood from younger and older adults, froze the samples under standard conditions, and then measured NAD+ after thawing, the measured values would reflect a combination of the true biological NAD+ concentration and the degree of freeze-thaw degradation each sample experienced. If, by chance or by systematic differences in sample handling, the older participants' samples experienced slightly more degradation than the younger participants' samples, the resulting data would show lower apparent NAD+ in older individuals. The statistical analysis would flag this as a significant age-related difference. The interpretation would be biological. But the cause would be a laboratory artifact.
The power analysis the Amsterdam UMC team performed makes the problem even more concrete. Under the variance structure typical of studies that include a single freeze-thaw cycle, detecting a genuine difference of 7 nanomoles per milliliter or less requires more than 20 participants per group. Several of the prior studies reporting age-related NAD+ declines in human blood used cohorts considerably smaller than this, and the effect sizes they reported were in many cases smaller than the threshold their sample sizes could reliably detect. This does not prove those studies were wrong in any specific instance. It does mean their findings were unlikely to be statistically robust even if the biological effect they were attempting to measure was real, and that handling-induced variability would have been sufficient to produce apparent effects of the magnitude they observed.
There is a further consideration worth naming directly. The studies that found no age-related NAD+ decline, which existed in the literature alongside those that did, may have been using sample handling procedures that better preserved NAD+ integrity, or working with cohorts where handling was more uniform, or simply operating in a regime where the noise was low enough that the absence of a signal was detectable. The inconsistency in the prior literature, which motivated this study in the first place, is itself consistent with a field where measurement methodology was driving results as much as biology was.
None of this means the researchers who conducted earlier studies were careless or that their findings were fabricated. Freeze-thaw degradation of NAD+ is a subtle and previously undercharacterized problem. Without the kind of systematic handling validation the Amsterdam UMC team performed, there was no obvious reason to suspect that standard sample storage practices were corrupting NAD+ measurements in a way that could produce spurious age-related differences. The field was operating with the tools and knowledge available to it. What this study provides is a clearer picture of what those tools were actually measuring, and what that means for conclusions drawn from them.
What This Does and Does Not Mean
A finding that challenges a widely held assumption requires careful interpretation, particularly when that assumption has become embedded in both the scientific literature and a commercially significant consumer market. The Amsterdam UMC study is a rigorous and important piece of work, and its conclusions deserve to be stated clearly. But so do its limits.
What this study does not say.
It does not say that NAD+ is unimportant for aging biology. The evidence that NAD+ plays a central role in mitochondrial function, DNA repair, sirtuin activity, and cellular stress response is substantial and largely derives from mechanistic work in cells and animal models that this study does not address. The biological case for NAD+ as a relevant molecule in aging remains intact.
It does not say that NAD+ levels do not decline with age in human tissues. The study measured whole blood, not muscle, liver, brain, or any other tissue where NAD+ metabolism matters most for the aging processes the supplement narrative is concerned with. It is entirely possible that tissue NAD+ declines with age in humans in ways that blood measurements cannot detect. Several studies using muscle biopsies have suggested that muscle NAD+ levels may be lower in older adults, and this study does not contradict those findings.
It does not say that NR or NMN supplementation produces no biological effects. The NR supplementation cohort in this study showed a clear increase in blood NAD+, confirming that direct precursor supplementation raises circulating NAD+ levels as expected. Whether that increase in blood NAD+ translates into meaningful increases in tissue NAD+ levels, and whether those tissue-level changes produce the functional benefits observed in animal models, are separate questions that this study was not designed to answer.
What this study does say.
Whole-blood NAD+ is not a reliable biomarker of biological aging in humans. Under rigorously controlled measurement conditions applied across seven independent cohorts totalling more than 300 individuals spanning a wide range of ages, fitness levels, and health statuses, blood NAD+ did not decline with age, did not differ between elite athletes and sedentary controls, and did not respond to exercise or nutritional interventions. The assumption that blood NAD+ tracks systemic aging has not held up under the most rigorous human testing yet applied to it.
This has a specific practical implication that deserves direct attention. Many individuals taking NAD+ supplements, and many researchers conducting NAD+ supplementation trials, have used blood NAD+ measurements as a readout of whether the intervention is working. The logic seems straightforward: if supplementation is raising blood NAD+, the intervention is engaging the target, and the biological benefits should follow. What this study reveals is that blood NAD+ was not the right readout to begin with, not because supplementation fails to raise it, it clearly does, but because blood NAD+ does not appear to reflect the age-related processes that supplementation is intended to address. Raising a biomarker that does not track the underlying biology is not the same as engaging that biology.
The tissue versus blood distinction is critical here and worth examining carefully. NAD+ is predominantly an intracellular molecule. The concentrations in whole blood, which this study measured, are driven primarily by the NAD+ content of red blood cells. Red blood cells are metabolically unusual: they lack mitochondria, do not perform oxidative phosphorylation, and are not subject to the same age-related metabolic changes that affect the tissues where NAD+-dependent processes like sirtuin activation and mitochondrial function are most relevant to aging. The NAD+ in red blood cells is doing something, but it may not be doing the same things that NAD+ does in muscle cells, neurons, or hepatocytes, and its level may not reflect what is happening in those tissues in any straightforward way.
This does not mean blood measurements are worthless for monitoring NAD+ supplementation. It means they should be interpreted for what they are: a confirmation that a supplement is being absorbed and converted to NAD+ in the bloodstream, not a readout of whether aging biology in relevant tissues is being meaningfully altered. Whether that tissue-level alteration is occurring, and whether it produces the functional benefits the supplement narrative promises, remains an open question that blood NAD+ measurements cannot answer.
What Comes Next
A study that removes a biomarker from the toolkit does not leave the field without options. It clarifies what the field actually needs and points toward where the harder but more important work lies.
The most immediate implication of this study is for the design of NAD+ supplementation trials in humans. If blood NAD+ is not a reliable readout of aging biology, then trials that use blood NAD+ as a primary endpoint are measuring the wrong thing. They can establish that a supplement raises blood NAD+, which this study confirms it does, but they cannot establish whether that elevation is meaningful for the aging processes the intervention is targeting. A trial that shows blood NAD+ increased by 50 percent following supplementation has demonstrated pharmacological activity. It has not demonstrated biological relevance in the tissues where that relevance would need to manifest.
What is needed instead are trials designed around tissue-level measurements and functional outcomes. Muscle biopsies can assess NAD+ concentrations in skeletal muscle, one of the tissues most relevant to the age-related decline in mitochondrial function that NAD+ supplementation is supposed to address. Functional readouts of mitochondrial efficiency, such as measurements of oxidative phosphorylation capacity or maximal oxygen consumption, can indicate whether tissue-level changes in NAD+ metabolism are producing the bioenergetic improvements the hypothesis predicts. Biomarkers of sirtuin activity and DNA repair efficiency could provide indirect evidence that NAD+-dependent cellular processes are responding to supplementation in ways that matter for aging biology. These measurements are more invasive, more expensive, and more difficult to conduct at scale than a blood draw, which is precisely why blood NAD+ became the default readout in the first place. But using an accessible measurement that does not track the biology is not a reasonable substitute for measuring the biology directly.
The broader lesson extends beyond NAD+ to the field of longevity biomarker research more generally. The gap between a compelling biological hypothesis and robust human evidence is consistently wider than it appears from animal model data, and the quality of measurement methodology plays a larger role in shaping apparent conclusions than is always acknowledged. The NAD+ story illustrates this pattern with particular clarity. A biologically coherent hypothesis, supported by compelling animal data and several human studies, generated enormous commercial momentum and research investment before the most basic methodological questions about how to measure the relevant biomarker correctly had been adequately answered. The result was a literature in which genuine biological signal and measurement artifact were intermingled in ways that could not be disentangled without the kind of systematic validation work the Amsterdam UMC team has now performed.
This is not a failure unique to NAD+ research. It is a recurring pattern in fields where the pressure to translate animal findings into human applications outpaces the development of rigorous human measurement tools. Epigenetic clocks, inflammatory biomarkers, telomere length, and a range of other aging-related measurements have all been subject to similar methodological debates about whether they are tracking biology or artifacts of the measurement process. The solution in each case is the same: invest in measurement validation before drawing conclusions from the measurements, and be willing to revise those conclusions when better methods reveal that earlier findings were not as robust as they appeared.
For the NAD+ field specifically, the path forward likely involves a combination of better tissue measurement approaches, more carefully controlled supplementation trials with functional endpoints, and a more honest accounting of what blood NAD+ measurements can and cannot tell us. The biological hypothesis underlying NAD+ research remains worth pursuing. The animal data is real, the cellular mechanisms are well characterized, and the possibility that NAD+ metabolism is a meaningful lever for human aging biology has not been ruled out by this study. What has been ruled out is the specific claim that blood NAD+ tracks aging in a way that makes it useful as a biomarker for monitoring that process or its reversal through supplementation.
The field is better off knowing this than not knowing it. A measurement that misleads is worse than no measurement at all, because it creates false confidence that the underlying biology is being monitored when it is not. The Amsterdam UMC study has done the field a genuine service by establishing this clearly, even if the clarity it provides is uncomfortable for a market that has been built on a different assumption.
Conclusion: What the Stability of Blood NAD+ Actually Tells Us
The finding at the center of this study is easy to misread. Blood NAD+ does not decline with age, does not respond to exercise, and does not respond to nutritional supplementation. For anyone who has been following the NAD+ narrative in longevity science, that result can feel like a deflation, a signal that the molecule everyone has been excited about is less important than believed. That reading would be a mistake.
What this study established is not that NAD+ biology is unimportant. It is that blood NAD+ is the wrong window through which to observe it. The molecule's role in mitochondrial energy production, sirtuin regulation, DNA repair, and cellular stress response is grounded in decades of mechanistic work that a single human biomarker study does not touch. The animal data showing that tissue NAD+ declines with age and that restoring it produces meaningful functional improvements remains intact. The biological case for NAD+ as a relevant target in aging research has not been weakened by this finding.
What has been weakened is the specific framework through which much of the human NAD+ research has been conducted. The assumption that blood NAD+ tracks systemic aging, that it declines with age in a way detectable through routine blood testing, and that raising it through supplementation provides a reliable signal that aging biology is being engaged, has not held up under rigorous measurement. Seven independent human cohorts, more than 300 individuals, a measurement method carefully validated against the handling artifacts that corrupted earlier work: the evidence is as clear as human research of this kind can produce.
For anyone currently taking NR or NMN supplements, this finding does not provide a reason to stop and does not provide a reason to continue. What it removes is one of the primary pieces of evidence that was being offered in support of supplementation as a longevity intervention in humans, specifically the claim that blood NAD+ declines with age and that measuring it before and after supplementation provides a meaningful readout of biological engagement. That claim was not supported by the most rigorous test it has yet received.
The supplement industry built significant commercial momentum around a narrative that was biologically coherent and supported by real animal data but that was translated into human applications before the foundational measurement questions had been answered. That is a familiar pattern in longevity science, where the distance between a compelling hypothesis and a validated human intervention is consistently longer than the excitement generated by early animal findings suggests. Navigating that distance honestly, acknowledging what the evidence does and does not support at each stage, is what responsible longevity science requires.
This study is an example of that navigation done well. The Amsterdam UMC team did not set out to debunk NAD+ research. They set out to measure it properly. What they found when they did should redirect the field toward better tools, better endpoints, and a more honest accounting of what human evidence for NAD+ interventions actually looks like. That redirection is not a setback for longevity science. It is how the field matures.
The blood was stable. The science is still asking the right questions. It just needs better ways to answer them.
- Trętowicz, M.M., Scantlebery, A.M.L., Schomakers, B.V. et al. Human whole-blood NAD+ levels do not vary with age or lifestyle interventions. Nat Metab (2026). https://doi.org/10.1038/s42255-026-01537-5
Related studies